
P. perezi

Rana atacada por un cangrejo rojo (Procambarus clarkii)
viernes, 30 de diciembre de 2011
domingo, 23 de octubre de 2011
Rana temporaria
 |
| Rana bermeja |
Origen: El género procede de Eurasia, existen restos Paleolíticos hace 15-20 millones de años; seguramente ya estaría presente en la Península Ibérica desde esas fechas, aunque sólo se han encontrado restos fósiles de 1.5 m.a.
Identificación: Se trata de una rana grande y robusta. Se distingue de la rana patilarga, además de por el tamaño, por poseer una serie de manchas marrones o negras en forma de V detrás de la cabeza, con el vértice dirigido hacía delante.
Tamaño: Los machos alcanzan los 30-40 mm y las hembras hasta 80-100 mm en Sanabria.
Descripción: Tienen hocico redondeado y cabeza corta. Los ojos son grandes y prominentes, muy separados entre si; la pupila es horizontal. Los tímpanos son grandes y visibles. Las extremidades posteriores, largas, están rematadas por cinco dedos unidos entre si por una membrana interdigital; los miembros anteriores son muy cortos (más que en otras ranas rojas), con cuatro dedos y sin membrana. La piel es lisa y puede presentar pequeñas rugosidades. La coloración es pardo-rojiza que con el tiempo puede adquirir tonalidades oliváceas, grisáceas o amarillas. La zona ventral es blanca o amarillenta.
Reproducción: La época de celo se inicia a partir de febrero hasta mayo, dependiendo de las zonas (en Galicia comienza en diciembre), en Sanabria se produce después del ciclo invernal con las primeras subidas de las temperaturas. Los machos llegan primero a las zonas de reproducción, atrayendo a las hembras mediante el canto muy estridente. La actividad sexual dura entre 15-40 días. El amplexus es axilar. La puesta se realiza con temperaturas mínimas de entre 4º a 8º C, y se compone de 900 a 4000 huevos depositados en masas globosas de cierto tamaño. Los huevos miden 3 mm y eclosionan después de 21-30 días, dependiendo de las temperaturas del agua. Los renacuajos alcanzan los 56 mm y los metamórficos unos 20-40 mm, su coloración es idéntica a la de los adultos. La metamorfosis varia de 65 a 115 días. La madurez sexual la alcanzan a los tres años.
Hábitat: Se limita a zonas de alta montaña, hasta 2.600 m.s.n.m., en entornos dominados por brezales, hayedos, robledales, pastizales y praderas de turba. En Sanabria se localiza básicamente en turberas de Peña Trevinca y en la Cabrera.
Alimentación: Se basa en Arácnidos, Coleópteros, Miriápodos, Colémbolos, Himenópteros, Ácaros, larvas de Dípteros y Anélidos. También se ha citado depredación sobre otras ranas en Galicia (Galán, 1993)
Depredadores: La introducción de salmónidos alóctonos como la trucha arco iris (Oncorhynchus mykiss) y el salvelino (Salvelinus fontanalis) han mermado nuestras poblaciones. La víbora de Seoane (Vipera Seoane) y la culebra de collar (N. natrix) y viperina (N. maura) fueron siempre sus depredadores naturales.
Observaciones: Está muy ligada a los medios acuáticos; es de costumbres nocturnas y diurnas indiferentemente. Pueden vivir diez años. La subespecie de Zamora es la Rana temporaria parvipalmata, que se caracteriza por tener menor extensión en la membranas interdigitales, la cabeza es más larga y estrecha; existe aún otra subespecie en el Pirineo y Navarra, la Rana temporaria temporaria, que presenta una menor longitud de las extremidades posteriores que la de Zamora.
 |
| Se han encontrado poblaciones en Sanabria por J. Alfredo Hernández y posteriormente Pedro Galán, la cuadricula más al sur debe ser descartada, puede tratarse de una confusión en la identificación . |
sábado, 24 de septiembre de 2011
Pelobates cultripes
 |
| Macho en Matalobas (Toro). Abel Bermejo. |
 |
| Hembra en Valdemantas(Valdefinjas). Abel Bermejo. |
Origen: Se conocen restos fósiles del Cretácico (100 m.a) en Norteamérica y Asía, del Eoceno Medio en Europa hasta el Mioceno (15 m.a), época en que ya presentaban una distribución circunmediterránea.
Identificación: Es un sapo de aspecto de rana, más rechoncho; carece de glándulas parotídeas.
Tamaño: La longitud de cabeza y cuerpo alcanza 110 mm en machos y 122 mm en hembras.
Descripción: Tiene cabeza ancha y morro redondeado, ligeramente puntiagudo. El tímpano no visible. Tiene algunas pequeñas verrugas glandulares en los costados. Su pupila es vertical y el iris plateado o verdoso. Presenta una fuerte coosoficación de la piel de la región cefálica dorsal (cráneo). La piel es lisa, con pequeñas verrugas en los costados. La coloración varía mucho, desde pardo jaspeado de tonos verdosos (conforman bandas longitudinales) a marrón oscuro con manchas patentes oscuras. La coloración ventral es blanca con algunas machas pardas o verdosas, especialmente en la garganta. No presenta sacos vocales. Las extremidades posteriores están adaptadas para el salto, con cinco dedos unidos por una membrana interdigital. Poseen una espuela metatarsal de color negro. Las extremidades anteriores son más pequeñas y robustas (especialmente en los machos) y presentas cuatro dedos. En esta zona se dan casos de ejemplares melánicos negros y de albinismo. Es nocturno.
Reproducción: Comienza en febrero y dura hasta mayo. Se han visto amplexus en diciembre y enero. El adelanto o retraso del celo depende sobre todo de las lluvias y la humedad. El amplexus es inguinal; la puesta se presenta en un grueso cordón de 1.500 a 4.000 huevos. El desarrollo embrionario dura de 14 a 16 días, los renacuajos son indistintamente vegetarianos y carnívoros. El periodo larvarios dura de 4 a 5 meses. Los metamórficos miden de 22 a 35 mm. La coloración es idéntica a la de los adultos y llevan una vida totalmente terrestre.
Hábitat: Aparece en zonas graníticas o calcáreas con cierta inclinación por los suelos sueltos, arenosos y arcillosos, siempre cerca de lagunas o charcas permanentes o estaciónales (rara vez se alejan de la masa de agua). Su forma de vida subterránea hace que la especie busque sustratos blandos donde existen formaciones de granito. Asciende hasta los 1.200 m.s.n.m.
Alimentación: Se alimenta de Arácnidos, larvas de insectos, Coleópteros, Anélidos, Gasterópodos, Himenópteros, Formícidos, Ortópteros, Hemípteros y polillas. Caza al acecho en caminos o zonas despejadas, eleva el cuerpo, quedándose inmóvil y a la espera de que pase algún invertebrado, momento en el que se abalanza sobre su presa.
Depredadores: Tiene muchos enemigos: el cangrejo rojo americano (Procambarus clarkii) es destructor de puestas y larvas, ofidios como la culebra viperina (Natrix maura) y de collar (Natrix maura), lechuza (Tyto alba), garza (Ardea cinerea), ratonero común (Buteo buteo), milano real (Milvus milvus), águila culebrera (Circaetus gallicus), cuervo (Corvux corax), mamíferos como la rata (Rattus norvegicus). Dentro de los anfibios se ha citado depredación sobre las larvas de esta especie por parte del gallipato (Pleurodeles waltl).
Observaciones: Solamente se encuentra activo en días muy húmedos y templados, por encima de 3.5ºC de temperatura y un 75-100% de humedad ambiental. Puede permanecer hasta dos años enterrado en espera de las lluvias. Se trata de una especie de distribución circunmediterránea. En invierno se entierra en galerías subterráneas de 6 a 20 centímetros de profundidad y 5 cm de diámetro, allí permanece en posición horizontal, con el cuerpo dirigido hacia el fondo, manteniendo el cráneo en la parte superior.
 |
| Especie bien distribuida, falta en algunas zonas del Oeste. |
Discoglossus galganoi
 |
| Ejemplar adulto. Iñigo Martínez-Solano |
Origen: Este género está compuesto por Discoglossus (único viviente) y Alytes; el sapo pintojo ibérico se separó de Alytes (sapos parteros) hace unos siete millones de años. Se conocen restos fósiles desde el Mioceno al Pleistoceno Medio en la península Ibérica. La separación del sapo pintojo ibérico de sus parientes más cercanos como D. pictus y D. sardus se produciría durante el Plioceno Medio hace unos tres millones de años. El sapillo pintojo ibérico es un endemismo de la Península.
Identificación: Tiene una pupila abombada muy perceptible, con aspecto de rana, con la que se puede confundir más que con los demás sapos. Los miembros posteriores largos, muy bien adaptados para el salto.
Tamaño: Es de tipo medio, en nuestra zona los machos tienen hasta 78 mm y las hembras 69 mm.
Descripción: Poseen una cabeza deprimida y ancha, con hocico puntiagudo y largo, similar a las ranas. El tímpano, poco marcado, es de menor tamaño que el ojo, cuyo iris es dorado. Los miembros anteriores son fuertes, con cuatro dedos cortos, puntiagudos, y desiguales, el primero es el más corto y el tercero el más largo, el segundo es algo más largo que el cuarto. Posee tubérculos palmares, resaltando el central. Las extremidades posteriores son largas, tienen cinco dedos palmeados y aplastados. La piel dorsal es lisa con algunas verrugas a la altura del ojo. Presenta un pliegue glandular poco desarrollado. La piel es lisa, con pequeñas verrugas dispersas por el dorso, especialmente en los machos. La coloración dorsal es predominantemente pardo-verdosa (en ejemplares de la zona), olivácea y gris. Posee una banda temporal oscura que va desde el hocico hasta el tímpano. En el dorso presenta manchas que a veces forman gruesas bandas longitudinales, la coloración rayada dibuja tres bandas longitudinales amarillentas que se unen en el hocico, un rasgo que las diferencia a primera vista de las ranas. El macho carece de saco vocal pero tiene palmeaduras en los dedos de los miembros traseros, en cambio las hembras y los juveniles carecen de estas membranas interdigitales.
Reproducción: Comienza el celo en primavera, para el cual prefieren las charcas temporales a las corrientes de agua. La cópula se realiza en el agua, mediante amplexus inguinal. La hembra deposita hasta 350-700 huevos, pero una misma hembra puede realizar varias puestas en la misma época (con varios machos), llegando en total a producir hasta 1.500 huevos en la reproducción anual. El diámetro exterior del huevo mide de 3-5 mm y la eclosión de los renacuajos se produce a los 5-10 días. En un primer momento las larvas miden de 3-4 mm, pero experimentan un rápido desarrollo, de manera que a las pocas horas de nacer pueden alcanzar ya de 6-7 mm; la medida máxima que alcanzan los renacuajos es de 36 mm. La metamorfosis dura entre 30-60 días. Al emerger del agua los metamórficos miden 7-16 mm.
Hábitat: Es una especie poco exigente del medio acuático, soportando distintas calidades y niveles de agua; se encuentra en bosques caducifolios, pinares húmedos, prados, sotos y riberas de arroyos y ríos, incluso en estepas cerealistas. Ocasionalmente se han observados en cuevas y minas abandonadas en Galicia, y en otros entornos generalmente no muy alejados del agua. Asciende hasta los 1900 m.s.n.m. en Sanabria (cabecera del Tera).
Alimentación: Su dieta comprende Coleópteros, Himenópteros (Formícidos principalmente), Arácnidos, Miriápodos, Dípteros y Anélidos.
Depredadores: Sus depredadores tradicionales son las culebras de agua (N. maura, N. natrix), nutria (Lutra lutra) y jineta (Genetta genetta), también la lechuza (Tyto alba) y otras rapaces nocturnas.
Observaciones: Esta especie está muy ligada a los medios acuáticos. Tiene aspecto de rana y puede confundirnos a la hora de su identificación; la observación del tímpano poco marcado en esta especie nos servirá como referencia para diferenciarlo de las verdaderas ranas, ya que en éstas el tímpano está bien visible y marcado. Llega a vivir 10 años.
 |
| La distribución es desigual, y las poblaciones esteparias de Tierra de Campos se encuentran en retroceso. |
viernes, 9 de septiembre de 2011
Pelodytes punctatus
 |
| Larva albina parcial. Xavier Rivera Mula |
 |
| Sapillo moteado al Sur del Duero. Abel Bermejo |
Origen: Se han encontrado restos fósiles en Europa en el Eoceno (50-40 m.a) siendo un grupo muy antiguo, con características de los anfibios más primitivos; en el Sur de la Península nos encontramos un pariente próximo, el sapillo moteado ibérico (Pelodytes ibericus)
Identificación: Tiene aspecto frágil y delicado, con cierta apariencia de rana. Sus miembros posteriores son largos, adaptados para el salto. Se puede confundir con el sapo partero, que es más corpulento que el sapillo moteado.
Tamaño: Es pequeño, los machos miden 35 mm y las hembras 50 mm.
Descripción: Posee una cabeza tan larga como ancha, con hocico redondeado. No tiene glándulas parotídeas. Los ojos son grandes, con pupila vertical elíptica. El tímpano es pequeño y bien visible. Sus miembros anteriores son cortos, con cuatro dedos alargados, ensanchándose en la punta. Presenta tres tubérculos palmares en los miembros anteriores. Las patas posteriores son largas, tienen cinco dedos alargados y ligeramente palmeados en su base, con tubérculos subarticales. El dorso es de color gris oliváceo, con manchas verdes y amarillas. La zona ventral es blanca. Se desplaza dando saltos y trepa muy bien, es nocturno.
Reproducción: El ciclo comienza en primavera, prefiriendo charcas temporales a aguas corrientes. La cópula se realiza en el agua, por medio de amplexus inguinal. Sus puestas son muy dispares, la hembra puede depositar desde 40 hasta 400 huevos de 1,5-2 mm de diámetro, que adhiere a las plantas acuáticas. Los renacuajos eclosionan entre los 3-18 días y miden de 3-4 mm. La metamorfosis dura de 45-90 días, los metamórficos, al emerger del agua , miden de 14 a 23 mm.
Alimentación: Su dieta esta compuesta de Coleópteros, Arácnidos, larvas de Lepidópteros, Hemípteros, Heterópteros y Dermápteros.
Hábitat: En Zamora asciende hasta los 1000 m.s.n.m. y lo encontramos principalmente en la estepa cerealista, asociado a charcas temporales y fuentes con pilones o abrevaderos. Prefiere sustratos de yeso, calizas y los suelos arcillosos de Tierra de Campos; al Sur del Duero también se encuentra en zonas arenosas.
Depredadores: Sus principales enemigos son las especies de peces alóctonas introducidas, como el pez sol (Leppomis gibbosus), el pez gato (Ameirus melas), la gambusia (Gambussia holbrooki) y el cangrejo rojo americano (Procambarus clarkii), que depredan sobre sus puestas, larvas y adultos. Además de los reptiles tradicionales, se han citado entre las aves depredadoras la lechuza (Tyto alba) y el cernícalo primilla (Falco naumani).
Observaciones: Son muy dependientes y están muy ligados a los medios acuáticos, especialmente estancados, limpios y de poca profundidad (abrevaderos de fuentes naturales). Puede vivir hasta 7 años.
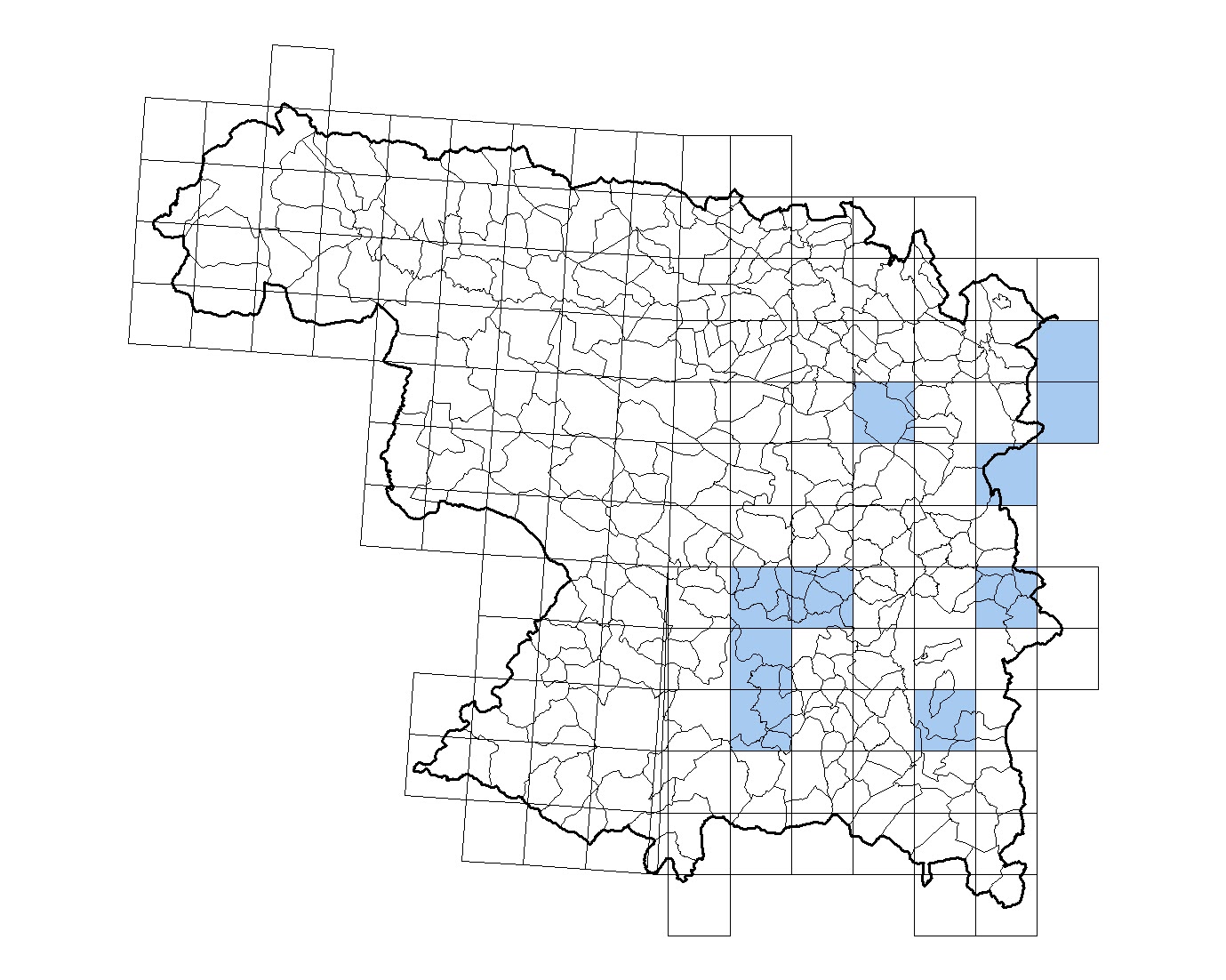 |
| Las poblaciones del norte están separadas de las del sur del Duero |
Alytes cisternasii
 |
| Macho con puesta. Iñigo Matínez-Solano |
 |
| Tras la eclosión, solo queda la envoltura. Abel Bermejo |
Origen: Los Alytes son un grupo muy antiguo con características de los anfibios primitivos, que aparecieron en el Cretácico Superior (90 m.a), el subgénero Ammorcyctis (sapo partero ibérico) se separó en el Mioceno Medio, hace 11-16 m.a.
Identificación: La piel presenta pequeñas verrugas. Pupila vertical, glándulas parotídeas poco patentes.
Tamaño: Sapo pequeño, alcanza un tamaño de 40 mm en los machos y 47 mm en las hembras.
Descripción: Tiene una cabeza ancha y pequeña, el hocico es corto, alto y redondeado. Los dedos carecen de tubérculos subarticulares. Su dorso es poco granuloso, salvo dos cordones glandulares correspondientes a las glándulas parotídeas (poco destacadas). El iris es dorado. La coloración del dorso es pardo-verdosa o gris salpicada por puntos anaranjado, el vientre tiene un color blanco sucio. Los machos son menores que las hembras, al contrario que en el partero común. Suelen presentar dos tubérculos palmares en las extremidades anteriores. No presentan membrana interdigital.
Reproducción: La cópula se realiza en tierra, el amplexus comienza siendo inguinal y finalmente pasa a ser cervical; el macho se sube a la espalda de la hembra y deja las extremidades posteriores debajo de la cloaca de ésta, luego comienza a realizar una serie de estiramientos de sus miembros posteriores (hasta 55 repeticiones), deteniéndose cuando la hembra adopta una posición encorvada y sitúa sus extremidades junto a las del macho, formando un receptáculo donde deposita los huevos. Una vez depositada la puesta, el macho realiza una serie de contracciones en sus flancos a la vez que los fecunda con su esperma. Luego, el macho suelta el amarre inguinal, que pasa a ser cervical; la hembra vuelve a adoptar la posición encogida hasta que los huevos quedan fijados en las extremidades posteriores del consorte, donde permanecen pegados por un cordón gelatinoso. Un macho puede transportar de 1 a 3 puestas de distintas hembras. El número de cada puesta suele variar entre 40-150 huevos, que miden 2,6 a 2,9 mm. El macho se encargará de mantenerlos húmedos e hidratados en todo momento. La puesta permanece 28 días con el macho , tras los cuales deposita los huevos ya maduros en el agua, de donde emergen los renacuajos (10-12mm) . Son también, como ocurre en las larvas del sapo partero común, vegetarianos y caníbales, indistintamente. La metamorfosis dura entre 3-9 meses. Los metamórficos miden de 12-25 mm.
Hábitat: Ocupa prados, dehesas con humedales, fuentes y abrevaderos rurales, bosquetes de robles y sotos frescos. Esta especie no coloniza altitudes tan elevadas como el sapo partero común, sólo hasta los 1, 200 metros de la sierra de la Culebra. Prefiere sustratos de suelos blandos de arenas (silíceas) y asociaciones graníticas en las inmediaciones, se puede observar ocasionalmente en suelos arcillosos en el Este de la provincia.
Alimentación: Se alimenta de Arácnidos, Coleópteros, Dípteros, Isópodos, Miriápodos, Dermápteros Himenópteros, Oligoquetos, Curculiónidos y en menor medida Anélidos.
Depredadores: Sus depredadores básicos son las culebras de agua (N. maura, N. natrix), y las rapaces nocturnas (lechuza).
Observaciones: Llegan a alcanzar los 8 años en cautividad. Es una especie eminentemente cavadora, realiza galerías subterráneas de hasta 20 centímetros de profundidad (datos de Zamora), en zonas arenosas entre masas graníticas (en Sayago), al contrario que el sapo partero común que prefiere refugiarse debajo de tocones, cortezas y piedras.
 |
| Esta presente en la zona Suroeste de la provincia y escasea hacia el Este de la provincia. Existen zonas donde se solapa con Alytes obstetricans . |
jueves, 8 de septiembre de 2011
Alytes obstretricans boscai
 |
| Macho acarreando la puesta en Toro (Zamora) |
 |
| La puesta puede constar de entre 20-160 huevos, que las hembras dan depositando en los machos. |
Origen: Los Alytes son un grupo muy antiguo que aparecieron en el Cretácico Superior (90 m.a), divergiendo del subgénero Ammorcyctis (sapo partero ibérico) en el Mioceno Medio, hace 11-16 m.a en la Península Ibérica, donde encontramos tres especies: sapo partero común (Alytes obstetricans), el único que habita en nuestra provincia, sapo partero balear (Alytes muletensis) y sapo partero bético (Alytes dickhilleni)
Identificación: La piel presenta pequeñas verrugas, pero mucho menos marcadas que en el sapo común o en el corredor. Tiene pupila vertical y glándulas parotídeas de forma no arriñonada, al contrario que en los anteriores. Poseen tres tubérculos palmares.
Tamaño: Es un sapo pequeño, los machos alcanzan 53 mm y 48 mm las hembras, que en esta especie, frente a lo que ocurre en la mayoría de los sapos, son menores.
Descripción: Poseen una cabeza más ancha que larga, con el hocico redondeado. Los dedos son cortos y aplastados (mucho más largo el central). Tiene el dorso cubierto de pequeñas verrugas lisas. El iris es dorado. La coloración es parda o gris, salpicada por manchas de tonos verdosos, las tonalidades son muy diferentes de una población a otra, no existen modelos fijos. Da grandes saltos con los miembros posteriores y es un excelente trepador.
Reproducción: La cópula se realiza en tierra, mediante amplexus inguinal, el macho fricciona repetidamente los flancos de la hembra hasta que ésta deposita los huevos sobre los miembros posteriores del macho a la vez que él los fecunda con su esperma. Los huevos quedan fijados al dorso y extremidades posteriores unidos por un cordón gelatinoso. En número de la puesta oscila entre 50-160 huevos. El macho se encargará de cuidar y mantener la puesta hidratada hasta el momento de la eclosión, llevándolos sobre su espalda. Permanecen de 19-21 días con el macho, hasta que éste, llegado el momento propicio, deposita la puesta en el agua, emergiendo los renacuajos (10-20 mm) poco después. Son vegetarianos y caníbales y suelen depredar sobre otras larvas de anfibios que comparten su medio, como el sapo de espuelas. La metamorfosis es larga y varia entre los 6 meses a los 2 años. Los jóvenes metamórficos miden de 15-30 mm al emerger del agua, y no se alejan mucho del punto de reproducción.
Hábitat: Prefiere sustratos rocosos o de tierra suelta (arenas y gravas), pero también está presente en sustratos silíceos, ácidos (pinares). En las zonas mesetárias tiene especial querencia por fuentes rústicas, pilones, abrevaderos y charcas permanentes. En zonas de media y alta montaña reside en bosques de roble melojo por debajo de 1.500 metros (Sanabria); en cotas superiores ocupa cabeceras de ríos, lagunas, lagos y turberas. Asciende hasta los 2,000 m.s.n.m en Zamora.
Alimentación: Su nutrición abarca Arácnidos, Fórmidos, Coleópteros, Dípteros, Isópodos, Colémbolos, Miriápodos y Anélidos.
Depredadores: Sobre la regresión de esta especie tienen especial incidencia las introducciones de salmónidos alóctonos en lagos y lagunas de montaña, igualmente el cangrejo americano y el galápago de orejas rojas; sus depredadores tradicionales son los mustélidos (Meles meles, Mustela putorius), culebras de agua (N. maura, N. natrix) y víboras (Vipera seoanei)
Observaciones: Es nocturno, activo desde febrero hasta octubre. Muy terrestre. Tiene dos celos, uno en primavera y otro en el otoño. Puede vivir 8 años. En lagunas de alta montaña, donde las estaciones favorables son de corta duración, las larvas tardan mucho tiempo en realizar la metamorfosis, llegando a permanecer hasta 8 años en fase larvaria, alcanzando entonces grandes tamaños de hasta 15-18 cm; curiosamente, cuando emergen estos sapos parteros el tamaño se reduce y son igual de pequeños que los metamorficos de otras poblaciones de menor altitud. La subespecie de Zamora es Alytes obstetricans boscai.
 |
| Esta bien asentado en la cuenca del río Duero y en el noroeste de la provincia. |
jueves, 1 de septiembre de 2011
Epidalea (bufo) calamita
 |
| Hembra en Toro. Abel Bermejo |
Origen: El sapo corredor se separó del sapo verde hace 20-25 m.a. colonizando Europa Occidental, hoy ocupa toda la Península Ibérica. Se conocen restos fósiles del Pleistoceno Medio ibérico y del Pleistoceno Superior en el sur de Portugal.
Identificación: Tiene, como el sapo común, pupila horizontal, y son muy perceptibles las glándulas parotídeas, paralelas entre sí. Sus extremidades son cortas, no adaptadas al salto, por eso camina, nunca salta. Su aspecto es rechoncho.
Tamaño: Es de tamaño medio, en la provincia los machos llegan a 95 mm y las hembras a 100 mm.
Descripción: Su cabeza más ancha que larga y el hocico corto, achatado. Tiene dedos cortos y puntiagudos, en los miembros anteriores el tercero es más largo; los dedos posteriores son cortos y aplastados (sin membranas interdigitales, ya que no es nadador). Tiene tubérculos palmares. La coloración es muy variable, predominando el verde grisáceo o con tonos pardos en el dorso; la zona ventral es blanco-amarillenta. Presenta una línea vertebral blanquecina o amarilla. No salta, se desplaza velozmente con rápidos movimientos de sus cortas extremidades (de ahí su nombre vulgar). Es una especie adaptada a la vida terrestre.
Reproducción: El celo comienza en primavera (dependiendo de la época de lluvias); en esa época coloniza lavajos de lluvia y charcas temporales de poca profundidad (10-30 cm) como lagunajos, charcos, cunetas y graveras regeneradas, siempre aguazales efímeros y someros. Suelen volver al mismo lugar todos los años. Los machos se reúnen en el agua antes que las hembras, emitiendo un canto muy sonoro (de ahí su nombre vernáculo), se han registrado grandes concentraciones de ejemplares machos (n=312) en los lugares de reproducción. La cópula se realiza en el agua, por medio de amplexus axilar. La hembra puede depositar hasta 10.000 huevos que el macho fertiliza montado en la espalda de la hembra, la fecundación es externa. Los embriones eclosionan del huevo a los 15 días, miden de 4-5 mm. La metamorfosis es muy rápida, en dos meses (incluso menos, pues aceleran el proceso si el agua es escasa); cuando emergen del agua miden 5,5-12,1 mm. Los metamorficos llevarán una vida terrestre y pueden alejarse mucho de la masa de agua donde crecieron.
Hábitat: Es amigo de zonas esteparias, campos de cultivo, pinares, graveras regeneradas; esta especie no requiere medios acuáticos permanentes y siempre que sea posible los evita empleando preferentemente charcas temporales. Asciende hasta los 1.800-2000 m.s.n.m. en Sanabria, ocupando turberas y lagunas de origen glacial.
Alimentación: Su dieta esencial la constituyen distintos tipos de Coleópteros, Arácnidos, Formícidos, Lepidópteros, Miriápodos, Gasterópodos y Anélidos.
Depredadores: Sus principales depredadores son los mustélidos (Meles meles y Mustela putorius), el jabalí (Sus scrofa) y la culebra de agua (N. maura); estos depredadores (salvo los ofidios) quitan la piel de este sapo antes de comerlo pues es muy tóxica.
Observaciones: Es muy terrestre (sólo acude al agua en la época de celo), avanza levantando el cuerpo y correteando, de ahí su nombre. En esta zona está activo desde febrero a noviembre. Popularmente se le conoce como sapo campanero debido a su atronador y sonoro canto, que invade corazón del aire en las noches primaverales, especialmente durante la época de reproducción; en algunas charcas se han llegado a contabilizar más de 300 machos cantando al unísono. Durante la hibernación se han observado ejemplares de sapo corredor cohabitando en las madrigueras del ratón de campo sin existir oposición de éste frente al sapo.
 |
| Típico desplazamiento de la especie (corredor). Abel Bermejo |
 |
| Al igual que Bufo bufo esta bien distribuido en la provincia, ocupa todo tipo de hábitat. |
Bufo bufo
 |
| Macho en Matalobas (Toro). Abel Bermejo. |
 |
| Hembra en Toro. Abel Bermejo |
Origen: Se conocen restos fósiles que datan del Paleoceno; el Bufo bufo se ha citado en el Pleistoceno medio Ibérico hace 17 m.a. Tiene relación genealógica con otras especies del género Bufo, como son el sapo verde (Pseudepiladea viridis) y el sapo corredor (Epidalea calamita); se separó evolutivamente de ambas hace 15 m.a.
Identificación: Tiene piel áspera con muchas verrugas. La pupila es horizontal. La coloración parda, levemente rojiza o jaspeada de pardo oscuro y amarillento.
Tamaño: Su longitud alcanza 115 mm en los machos y 190 mm en las hembras. Hemos hallado hembras viejas en la zona de Toro que llegan a alcanzar los 310 mm.
Descripción: Tienen la cabeza más larga que ancha, con hocico corto y redondeado. La pupila de horizontal es un rasgo específico. Posee glándulas parotídeas muy marcadas y oblicuas entre sí. Los dedos de las extremidades anteriores son cortos, y el tercero es más largo que el resto; en los miembros posteriores esos dedos son más alargados y aplastados y poseen membrana interdigital. Se desplaza dando saltos y es buen nadador.
Reproducción: Su época de celo tiene lugar entre febrero y mayo; para aparearse suelen visitar la misma charca todos los años. Él número de machos es muy superior al de las hembras, pudiéndose ver durante el celo a varios machos (n=6) encima de una sola hembra. El amplexus es axilar. La puesta puede constar de 1.500 a 4.000 huevos. El embrión se desarrolla en 2-3 semanas, los renacuajos son vegetarianos y tardan de 70 a 120 días. Los metamórficos miden entre 7,5-14 mm, llevarán una vida terrestre y no retornarán a la charca hasta que sean adultos (3-4 años).
Hábitat: Son extraordinariamente terrestres, se les encuentra en pinares, sotos con bosques de ribera, prados, brezales, cerca de lagunas de montaña (Sanabria), estepas y campos de cultivo, etc., aunque siempre asociados a un punto acuático permanente. Asciende hasta los 1800-2000 m.s.n.m. en zonas de alta montaña, en Sanabria.
Alimentación: Es muy voraz y come grandes cantidades de todo tipo de Coleópteros, Formícidos, Ortópteros, Arácnidos, Miriápodos, Anélidos, incluso otros anfibios y hasta pequeños mamíferos (roedores).
Depredadores: Son presas frecuentes de las culebras de agua (N. maura y N. natrix), nutria (Lutra lutra), cigüeñas (C. Ciconia y C. nigra), garzas (A. cinerea y A. purpúrea)), ratonero (Buteo buteo), milano negro (Milvus migrans), águila culebrera (Circaetus gallicus), etc.; otro problema nuevo es la introducción del cangrejo rojo americano (Procambarus clarkii) y de peces depredadores, como el pez sol (Leppomis gibbosus), que destruyen los puestas y larvas.
Observaciones: Pasa un periodo de hibernación desde noviembre a febrero; aunque es nocturno se nuestra activo en los días lluviosos. Es una especie muy terrestre que se aleja mucho del agua.
Existen tres subespecies de Bufo, B. b. spinosus, B. b. bufo y B. b. gredosicola
 |
| Es una especie que se encuentra bien distribuida en Zamora, pero en los últimos años sus poblaciones se han visto reducidas. |
viernes, 26 de agosto de 2011
Lissotriton helveticus
 |
| Metamórfico. Abel Bermejo |
 |
| Macho adulto en fase acuática. Abel Bermejo |
Origen: Al igual que Lissotriton boscai esta especie se separa del género Triturus hace 6-7 millones de años, aparecen restos fósiles en Europa central donde evolucionará. En la Península Ibérica quedó aislado y separado genéticamente cuando el paso de los Pirineos se cerró.
Identificación: Se trata de un tritón de pequeño tamaño. Posee una banda a cada lado de la cabeza a modo de antifaz, y una franja amarilla en el vientre.
Tamaño: Los machos pueden llegar a los 85 mm y 90 mm las hembras.
Descripción: Tiene la cabeza más larga que ancha. Presenta tres surcos longitudinales en la parte superior de la cabeza. Los machos presentan cresta dorsal, las hembras no desarrollan crestas. Las colas de ambas sexos son diferentes, en el macho rematada en forma de lanza, en la hembra la punta es roma. Los machos muestran palmeaduras en sus extremidades posteriores de color oscuro en el celo. El color dorsal es pardo claro, el ventral tiene un ancha franja amarillenta.
Reproducción: En Sanabria el ciclo reproductor va desde enero hasta abril. Los cortejos del tritón palmeado son similares a las otras especies del género Triturus, esto es, se basa en movimientos ondulatorios de su cola por parte del macho para provocar la reproducción. La época de freza (puesta) se puede extender hasta mayo, depositando de 300-500 huevos. El desarrollo embrionario dura 18-23 días. Los metamórficos miden de 26-35 mm.
Hábitat: Ocupa medios con cierta vegetación (especialmente elige para la puesta el género de plantas conocidas como Chara sp.); no es exigente con el sustrato (calizos, graníticos y silíceos) ni con el medio o entorno vegetal: robledales, castañales, turberas, estanques artificiales, arquetas de riego, fuentes y pilones... Está presente en zonas de una pluviometría superior a los 700 mm anuales. Asciende hasta los 2.500 m.s.n.m.
Alimentación: Se alimenta de larvas de Coleópteros, ninfas, Dípteros, Isópodos, Odonatos, Plecópteros, Dermápteros, Ostrácodos, Gasterópodos y Anélidos.
Depredadores: Sus enemigos tradicionales son la culebra de agua (Natrix maura) y las víboras (Vipera seonaei), pero actualmente el mayor peligro reside en las introducciones de salmónidos en zonas de reproducción.
Observaciones: En esta especie se da el fenómeno de la “neotenia” (ya culminada la metamorfosis mantienen aún durante un tiempo las branquias respiratorias) en una parte de las larvas, completándose al año siguiente. Existen tres subespecies: L. helveticus alonsoi, L. helveticus helveticus y L. helveticus punctillatus.
 |
| Solamente existe una población en Sanabria, es la especie más amenazada de Zamora. Recientemente se han encontrado ejemplares de la especie en la cuadricula 10x10 PG77. |
Lissotriton boscai
 |
| Macho en fase acuática. Abel Bermejo |
 |
| Hembra melánica grávida. Abel Bermejo |
Origen: El género Lissotriton aparece en el Eoceno-Oligoceno, momento en que se separa del género Triturus, hace unos 6-7 millones de años. Esta especie evolucionará de forma aislada en la Península Ibérica, se trata, pues, de un endemismo, como expresa su nombre.
Identificación: Presenta en el dorso una coloración pardusca, preferentemente verde oliva con manchas negras, en cambio el vientre es anaranjado o rojizo con manchas oscuras de forma redondeada. En algunas zonas la larga permanencia en el agua oscurece la coloración, al igual que en la fase terrestre.
Tamaño: Es el tritón más pequeño, la longitud total en machos ronda los 85 mm y los 96 mm en hembras.
Descripción: Tiene la cabeza tan larga como ancha, con el hocico redondeado. Los dedos no presentan membranas interdigitales. Se aprecia un surco longitudinal en la parte inferior de la cabeza. Las glándulas parotídeas son muy patentes. La longitud de la cola es igual o mayor que la cabeza y cuerpo juntos. Tiene cola con crestas poco desarrolladas, terminadas en punta, pero carece de cresta dorsal. La coloración varía desde el pardo oliváceo, al pardo verdoso, el vientre es anaranjado, amarillento o rojizo, con manchas negras grandes y de forma redondeada, se han descrito ejemplares con el vientre blanco.
Reproducción: El periodo de celo en nuestra provincia va desde octubre hasta mayo; después del cortejo el macho deposita el espermatóforo (4-5 mm) en el fondo del medio acuático, seguidamente lleva a la hembra hacía el mismo para que esta lo recoja con su cloaca. La fecundación es interna, días después comenzará a depositar entre 150-300 huevos. Las larvas alcanzan los 18 mm. La metamorfosis dura de 80 a 20 días.
Hábitat: Es frecuente en fuentes, pilones, acequias y charcas de aguas quietas. Precisa de aguas limpias y sin contaminación, tolera pH ácidos y alcalinos, y se adapta a sustratos graníticos, arcillosos, arenosos y fangosos. Asciende hasta los 1.200 m.s.n.m.
Alimentación: Come larvas de Coleópteros, Ninfas, Dípteros, Crustáceos, Ostrácodos y Anélidos. Se ha citado el consumo de puestas de sapo partero común.
Depredadores: Su mayor peligro lo representan especies alóctonas como el cangrejo rojo americano y los peces introducidos en las zonas de freza (pez sol, carpín dorado); también, pero en menos medida, la culebra de agua (N. maura, N. natrix), la cigüeña (Ciconia ciconia y Ciconia nigra), la garza (Ardea cinerea Ardea purpurea), la lechuza (Tyto alba) y mustélidos (Meles meles y Lutra lutra)
Observaciones: Es muy acuático, puede pasar todo el año en el agua. Se conocen ejemplares que han llegado a vivir 10 años en cautividad. Reduce la actividad por debajo de 2-4º C. Existen cinco grupos diferenciados entre si en la Península Ibérica.
 |
| Las poblaciones del Este debemos de darlas por extintas. |
jueves, 18 de agosto de 2011
Trabajos publicados por Abel Bermejo García
SEGUIMIENTO DE UNA COMUNIDAD DE ANFIBIOS A PARTIR DE LA CREACIÓN DE UNA CHARCA, EN TORO(ZAMORA)
Abel Bermejo García
C/ Cortes de Toro, 11, 2º,B. 49800 Toro. Zamora. España.
e-mail: gallipato@herpetologica.org
Keywords: monitoring , amphibian community, creation small pond, Toro, Zamora, Spain.
En los últimos años el declive poblacional de ciertas especies de anfibios ha llamado la atención de la sociedad en general, ya que han visto escasear o desaparecer especies de sapos o ranas que no hace mucho tiempo eran abundantes. La destrucción del hábitat, la contaminación, las enfermedades emergentes y la introducción de especies alóctonas son las principales causas de este declive(Barbadillo & García-París, 1991; Pleguezuelos, 1997; Montori et al., 2001; Cruz & Rebelo, 2005; Cruz et al 2006 ; Bermejo-García 2006). Los cambios en la agricultura, y con ello la desaparición de las balsas de riego que almacenaban agua para utilizarla en el estío, y el abandono del pastoreo extensivo ha provocado la perdida de abrevaderos tradicionales, desapareciendo uno de los biotopos más importantes para muchas especies de anfibios. La nula o mala conservación de estas fuentes ancestrales genera la pérdida de poblaciones establecidas en ese hábitat desde hace mucho tiempo. La sobreexplotación de los acuíferos en lugares que tradicionalmente se dedicaron al secano, está alterando el nivel freático, afectando a los manantiales que alimentan pilones, abrevaderos y fuentes. Como consecuencia de esto el aislamiento entre las masas de agua es un hecho y la posibilidad de contacto entre diferentes poblaciones que aún resisten en zonas aisladas es poco seguro debido a la gran distancia que las separa, a lo que hay que añadir el incremento de las vías de comunicación que producen aun mayor aislamiento. Con estas premisas, una de las mejores soluciones para evitar la desaparición de los anfibios en lugares donde en el pasado fueron abundantes es la creación de lagunas o charcas. Experiencias de otros autores confirman la rápida colonización de las nuevas masas de agua.(Reques. R. & Tejedo. M. 2008)
METODOLOGIA
Con esta finalidad, en 2002, se construyó una charca en un terreno particular, situado en el municipio de Toro (Zamora). Este terreno se había mantenido durante largo tiempo sin cultivos (15 años). La extensión total de la parcela que acoge la charca asciende a dos hectáreas, formadas por herbáceas y arbustos de pequeño porte permanentes, cantueso (Lavandula stoechas), ruda (Salvietum lavandulifoliae) y tomillo (Thymus spp), y temporales como cebadilla ratonera (Hordeum murinum), amapola (Papaver rhoeas), malva (Malva silvestris) y zurrón de pastor (Capsella bursa-pastoris) entre otras. Está situada en la cuenca sedimentaria del río Duero a una altitud de 689 metros. El suelo está formado por depósitos aluviales de distinta granulometría y arcillas que le dan consistencia.
En el lugar se había detectado la presencia de varias especies de anfibios antes de crear la laguna: Pelobates cultripes, Alytes obstetricans, Bufo calamita, Bufo bufo y Pelophylax perezi.
Esta presencia se debe a la existencia de dos fuentes cercanas, que a pesar de las adversidades todavía mantenían agua, en 2002 (hoy están definitivamente secas), lo que permitía la reproducción de algunas especies. Valorando la presencia de estos anfibios se perfiló el tamaño de la masa de agua y la creación de varios ambientes: fuente, pilones, charca y muros de protección, para atraer al mayor número de especies. Los cálculos finales dieron lugar a una pequeña laguna de 40 metros cuadrados y 80 cm de profundidad máxima, con una capacidad aproximada de 60.000 litros de agua. Se construyó un murete en la parte norte del estanque para protegerlo del viento del noreste en invierno. Se creó otra estructura que alberga un bidón de plástico de 200 litros, con arenas y cantos rodados para el decantado del agua El sistema funciona con una pequeña bomba que impulsa el agua al bidón y de éste cae al estanque. Más tarde se decidió construir una fuente tradicional techada con un arco de medio punto, y aljibe de agua que vierte a dos pilones que a su vez desaguan de nuevo en el estanque. El llenado se realiza mensualmente en verano y se emplea agua de pozo. El diseño es típico de la zona, pues pudimos comprobar que encontrábamos más especies de anfibios en lugares donde estaba presente el conjunto de estas construcciones, es decir, fuente, pilones y charca, comparándolos con parajes que sólo presentaban pilones o pilones con charca, o charca solitaria (datos propios). Se pudo constatar que la ocupación de los pilones por el sapo partero, respecto a otras especies, es mayor, y es el único en colonizar estas construcciones artificiales de difícil acceso para otros anfibios (Barbadillo et al 1999; Velasco et al 2005; datos propios). En lugares donde existía una charca o laguna las probabilidades de encontrar varias especies aumentaba, especialmente en el caso de los urodelos debido a la mayor capacidad de agua y espacio (Salvador & García- París, 2001; Montori, A. & Herrero, P.2004).
Por otra parte, para evitar la aparición de enfermedades emergentes como la quitridiomicosis, se tuvieron en consideración las recomendaciones del Dr. Jaime Bosch (comunicación, 2006) del Museo de Ciencias Naturales de Madrid, que indican que en los lugares soleados y libres de obstáculos que pudieran proyectar sombra a la masa de agua, el quitridio no se manifiesta (puede estar latente), debido a un rango de temperatura constante que impide su desarrollo (temperaturas superiores a los 28ºC durante las estaciones más calurosas), y destruye las esporas del mismo. Estos datos se corresponden a zonas estudiadas en la meseta, donde la altitud media es de 700 metros sobre el nivel del mar. Valorando estas medidas, la situación del enclave es sur-oeste y la exposición solar es total en el trascurso del día. Esto, mantiene la charca en temperaturas subóptimas para el quitridio. Desde la creación de este entorno no se han producido mortalidades masivas en anfibios producidas por el hongo, a pesar de haberse detectado el quitridio en dos charcas cercanas (150 metros) afectando a Pelophilax perezi y Alytes obstetricans (Bosch, com. per.).
RESULTADOS
En febrero 2002 se realizó el llenado de laguna y tres meses después observamos la presencia de Pelophylax perezi (n=6.090)*. Con fecha de 06/06/2002 encontramos 6 machos adultos y 9 hembras de diversos tamaños, además ese mismo año se reproduce. Alytes obstetricans (n=3.956) se observó en septiembre de 2002, época en que se contabilizaron dos machos y dos hembras adultas reproductoras, y en octubre se localizaron larvas de pequeño tamaño en un número aproximado de 120. En cuanto a Pelobates cultripes (n=1.831) las observaciones se realizaron en abril de 2002, una pareja adulta en fase reproductora en el agua. A raíz de estas citas nos disponemos a hacer un recuento de la población año tras año hasta 2008. Se efectuaron censos cuatro veces por año (primavera-otoño) con las sumas totales de adultos, larvas y metamorficos, constatando un radical y progresivo aumento de las poblaciones de todas esas especies. Tabla 1
A estas especies se les va ha sumar Bufo calamita (n=581) en 2003, que se ha reproducido pero con gran variabilidad numérica en estos años, siendo un reproductor ocasional. La profundidad y la permanencia de agua de la charca no beneficia la presencia de este anuro. Bufo bufo (n=11) se ha censado pero sin reproducción alguna, a pesar de lo cual desde 2004 ha estado presente. Triturus marmoratus (n=24) fue observado en 2007 y 2008, y se hallaron metamórficos esta última temporada. Respecto a Pleurodeles waltl se ha observado un solo ejemplar macho adulto en 2008. El tamaño y la temporalidad de la masa de agua determina el número de especies que se reproducen en ella (Richter-Boix et al, 2006; Richter-Boix et al, 2007), siendo estos factores suficientes para albergar depredadores de anfibios, como la culebra viperina (Natrix maura) presente desde 2007. Carecemos de datos de reproducción de esta especie, pero la observación de machos y hembras (n=4) hace pensar que es posible. La presencia de este ofidio, pone de manifiesto la buena salud ecológica de que goza esta charca. En seis años de seguimiento hemos contabilizado 12.494 ejemplares (larvas y adultos) de las especies citadas.
CONCLUSIONES
Pensamos que iniciativas de este tipo ponen de manifiesto la necesidad de realizar proyectos simples y locales para no perder la rica y variada diversidad de anfibios y reptiles que tenemos en la Península Ibérica.
Creemos que instalar estanques artificiales en zonas privadas o publicas puede ayudar mucho a mejorar la biodiversidad batracológica y los contingentes poblacionales de los hábitats naturales fuera de lugares protegidos, sobre todo, cuando las fuentes y pequeñas charcas donde tradicionalmente han vivido desaparecen, por las razones antes expuestas. Este fenómeno nos alerta de la delicada situación de los humedales menores, y lo sencillo y eficaz que resultan para la conservación de anfibios este tipo de iniciativas particulares. El resultado es evidente, esta pequeña construcción ha posibilitado la permanencia de no menos de siete especies de anfibios y una de ofidio.
* número de ejemplares marcados en seis temporadas
Bibliografía:
Barbadillo, L.J. & García- París, M. 1991. Problemas de conservación de los anfibios de España. Quercus, 62: 12-16.
Barbadillo L. J., Lacomba, J. I., Pérez –Melado. V., Sancho. V & López –Jurado. L. F. 1999. Anfibios y Reptiles de la Península Ibérica; Baleares y Canarias. ED. Geoplaneta. Barcelona.
Bermejo-García, A. 2006. Primeros datos sobre agresiones de Procambarus clarkii sobre Pleurodeles waltl. Bol. Herpetol. Esp. (2006): 82-84.
Bosch, J. 2006. Aparición del hongo patógeno de anfibios Bratrachochytrium dendrobatidis en Europa: ¿introducción humana o consecuencia del calentamiento global?. IX Congreso Luso-Español-XIII Congreso Español de Herpetología, San Sebastián.17.
Cruz, M. J. & Rebelo, R. 2005. Vulnerabiliy of southwest Iberian amphibians to an crayfish, Procambarus clarkii. Amphibia-Reptilia, 26: 293-303.
Cruz, M, J., Rebelo, R., & Crespo, E. G. 2006. Predation by an exotic grayfish, Procambarus clarkii, on Natterjack Toad, Bufo calamita, embryos: its role on the exclusion of this amphibian from its breeding ponds. Copeia, 2006: 274-280.
Montori, A. y Herrero, P. 2004. Caudata. En; Amphibia, Lissamphibia. García-París, M. Montori, A. y Herrero, P. Fauna Ibérica. Vol. 24. Ramos M. A. et al. (eds). Museo Nacional de Ciencias Naturales. CSIC. Madrid: 43-275.
Montori, A,. Llorente, G.A., Carretero, M.A. & Santos, X. 2001. La gestión forestal en relación con la herpetofauna. In: Conservación de la biodiversidad y gestión forestal: su aplicación en la fauna vertebrada (Camprodon J. & Plana, E. eds.) Edición de la Universidad de Barcelona, Barcelona: 251-289.
Pleguezuelos, J.M. (ed) 1997. Distribución y biogeografía de los anfibios y reptiles de España y Portugal. Universidad de Granada y Asociación Herpetológica Española. Granada, 542 pp.
Reques, R & Tejedo, M .2008. Crear charcas para anfibios: Una herramienta eficaz de conservación. Quercus, 273: 14-20.
Richter-Boix, A., Llorente, G.A. & Montori, A. 2006. Breeding phenology of an amphibian community in Mediterranean area. Amphibian-Reptilia, 27: 549-559.
Salvador, A. & García-París, M. 2001. Anfibios Españoles. Canseco Editores, S.L. Talavera de la Reina.
Velasco, J.C., Lizana-Avía, M., Román-Sancho, J., Delibes de Castro, M., & Fernández-Gutiérrez, J. 2005. Guía de Peces, Anfibios, Reptiles y Mamíferos de Castilla y León. Nayalde Editorial. Medina del Campo. Valladolid.
Temporada | Especies | Machos | Hembras | Larvas | Totales | |
2002 | Pelophylax perezi Alytes obstetricans Pelobates cultripes | 6 2 1 | 9 2 1 | 367 120 169 | 382 124 171 | |
677 | ||||||
2003 | Pelophylax perezi Alytes obstetricans Pelobates cultripes Bufo calamita | 10 4 1 2 | 12 5 1 1 | 459 298 201 69 | 481 307 203 72 | |
1.063 | ||||||
2004 | Pelophylax perezi Alytes obstetricans Pelobates cultripes Bufo calamita Bufo bufo | 24 7 1 1 2 | 12 6 1 1 - | 671 401 198 168 - | 707 414 200 170 2 | |
1.493 | ||||||
2005 | Pelophylax perezi Alytes obstetricans Pelobates cultripes Bufo calamita Bufo bufo | 38 10 1 1 2 | 9 10 1 - - | 853 560 98 - - | 900 580 100 1 2 | |
1.583 | ||||||
2006 | Pelophylax perezi Alytes obstetricans Pelobates cultripes Bufo calamita Bufo bufo | 35 11 1 - 1 | 16 10 1 - - | 1.014 671 101 - - | 1.065 692 103 - 1 | |
1.861 | ||||||
2007 | Pelophylax perezi Alytes obstetricans Pelobates cultripes Bufo calamita Bufo bufo Triturus marmoratus Natrix maura | 42 14 2 2 2 1 - | 26 11 2 1 1 1 - | 1.112 789 361 333 - - - | 1.180 814 366 336 3 2 1 | |
2.702 | ||||||
2008 | Pelophylax perezi Alytes obstetricans Pelobates cultripes Bufo calamita Bufo bufo Triturus marmoratus Pleurodeles waltl Natrix maura | 46 16 2 2 2 3 1 2 | 25 10 2 - 1 3 - 1 | 1.304 998 681 - - 16 - - | 1.375 1.024 685 2 3 22 1 3 | |
3.115 | ||||||
12.494 |
Tabla1. Número de especies y el censo de adultos, larvas de las mismas desde 2002 a 2008
 |
| Juvenil de P. clarkii devorado por un ejemplar de mayor tamaño. Abel Bermejo |
- Bermejo-García, A. (2003): Primeros datos de agresiones de cangrejo rojo americano (Procambarus clarkii), sobre gallipato (Pleurodeles waltl). Butll. Soc. Cat. Herp. 15: 87-91
-Bermejo-García, A (2004): Estado de conservación de las poblaciones de Triturus marmoratus en la comarca de Toro(Zamora). Bol. Asoc. Herpetol. Esp. 15(1): 50-53
-Bermejo-García, A (2006ª) Impacto de la fauna exótica sobre los anfibios de Castilla y León. Quercus, 239: 30-31.
-Bermejo-García, A (2006b) Nuevos datos de agresiones de Procambarus clarkii sobre Pleurodeles waltl. Bol. Asoc. Herpetol. Esp. 17(2): 82-85
ESTADO DE CONSERVACIÓN DE LAS POBLACIONES DE TRITURUS MARMORATUS EN LA COMARCA DE TORO(ZAMORA)
Abel Bermejo García
C/ Cortes de Toro nº11, 2B, 48900-Toro, Zamora España
e-mail: herpeto31@hotmail.com; correopangea@ya.com
Key words: Monitoring, conservation, Triturus marmoratus, Toro district, Zamora province, NW Spain.
Actualmente la comarca de Toro alberga dos poblaciones donde se reproduce Triturus marmoratus, la población principal está al sur del Duero (UTM 30T TL8885), tratándose de una fuente manantial (fuente del Macho) la cual se emplea para regar los huertos colindantes. La otra población se sitúa al norte del Duero (UTM 30T TM9704). En este caso se trata de un arroyo intermitente llamado Adalia, en un tramo de 800 m donde el agua es permanente. La distancia entre ambas poblaciones es de 35 km aproximadamente. La población sur no ha sido colonizada por Procambarus clarkii ni peces, debido a su aislamiento. En cambio, la norte se ve afectada por introducciones deliberadas de peces, Cyprinus carpio y Rutilus arcasii, siendo esta última una especie autóctona que parece llevar tiempo en la zona. Se han observado ejemplares de Procambarus clarkii en algunas ocasiones pero no en gran numero, siendo introducciones esporádicas y no llegando a reproducirse.
La observación de las poblaciones comenzó en 1998 realizándose un censo de Triturus marmoratus, identificando adultos, juveniles, metamórficos y larvas. El marcado se realizó mediante amputación de dedos en adultos y juveniles y corte de cola en larvas. Para censar los peces se practicaron diversas prospecciones con redes en verano, cuando el arroyo quedaba reducido a un charco. Tanto el marcado del urodelo como el de los peces se lleva a cabo todos los años. Se realizaron unas 200 salidas aproximadamente en diferentes épocas del año.
Población | Observación | Nº de ejemplares | Especie |
NORTE | 1998 1999 2000 2001 2002 1998 1999 2000 2001 2002 | 220 260 323 336 452 2 4 5 5 5 | Rutilus arcasii" " " " Cyprinus carpio " " " |
Tabla 1. Censo de peces avistados desde el 1998.
Población | Observación | Nº de ejemplares | |
SurNorte | 1998 1999 2000 2001 2002 1998 1999 2000 2001 2002 | 580 521 688 878 1523 625 83 52 21 0 | |
Total 4971 |
Tabla 2. Avistamientos de individuos metamórficos de Triturus marmoratus.
Año | clase | Población norte | % | Fecha observación | Población sur | % | Fecha observación | total |
1998 | Hembras | 4 | 22.2 | 04/03/1998 | 11 | 61.1 | 15/05/1998 | 18 |
- | - | - | 1 | 5.5 | 22/07/1998 | |||
Machos | 4 | 44.4 | 02/03/1998 | 3 | 33.3 | 11/04/1998 | 9 | |
1 | 11.1 | 13/10/1998 | 1 | 11.1 | 30/10/1998 | |||
Juveniles | 2 | 25.0 | 30/08/1998 | 4 | 50.0 | 22/10/1998 | 8 | |
2 | 25.0 | 01/11/1998 | - | - | - | |||
Metamórficos | 625 | 51.8 | 25/05/1998 | 580 | 48.1 | 12/06/1998 | 1205 | |
Larvas | 711 | 53.7 | 10/04/1998 | 612 | 46.2 | 30/05/1998 | 1323 | |
5 | 0.37 | 18/05/1998 | - | - | - | |||
Total | 1354 | 52.8 | - | 1212 | 47.2 | - | 2563 | |
1999 | Hembras | 9 | 40.9 | 03/04/1999 | 13 | 59.0 | 12/03/1999 | 22 |
1 | 4.5 | 05/04/1999 | - | - | - | |||
Machos | 2 | 18.1 | 03/03/1999 | 6 | 54.5 | 05/03/1999 | 11 | |
2 | 18.1 | 22/10/1999 | 1 | 9.0 | 12/10/1999 | |||
Juveniles | 1 | 6.6 | 16/05/1999 | 2 | 13.3 | 22/07/1999 | 15 | |
2 | 13.3 | 17/05/1999 | 10 | 66.6 | 20/10/1999 | |||
Metamórficos | 83 | 12.9 | 30/06/1999 | 521 | 81.4 | 30/05/1999 | 640 | |
Larvas | - | - | - | - | - | - | 624 | |
- | - | - | 624 | 100 | 22/04/1999 | |||
Total | 100 | 7.8 | - | 1177 | 92.1 | - | 1277 | |
2000 | Hembras | 2 | 11.7 | 10/04/2000 | 14 | 82.3 | 21/03/2000 | 17 |
- | - | - | 1 | 5.8 | 01/06/2000 | |||
Machos | 1 | 11.1 | 07/03/2000 | 7 | 77.7 | 05/02/2000 | 9 | |
- | - | - | 1 | 11.1 | 11/10/2000 | |||
Juveniles | 1 | 7.1 | 12/06/2000 | 10 | 71.4 | 10/05/2000 | 14 | |
1 | 7.1 | 13/06/2000 | 2 | 14.2 | 20/05/2000 | |||
Metamórficos | 52 | 7.0 | 22/05/2000 | 688 | 92.9 | 02/05/2000 | 740 | |
Larvas | - | - | - | - | - | - | 521 | |
- | - | - | 521 | 100 | 22/02/2000 | |||
Total | 57 | 4.3 | - | 1244 | 95.6 | - | 1301 | |
2001 | Hembras | - | - | - | 18 | 78.2 | 09/03/2001 | 23 |
- | - | - | 5 | 21.7 | 10/10/2001 | |||
Machos | 1 | 16.6 | 20/05/2001 | 4 | 66.6 | 03/02/2001 | 6 | |
1 | 16.6 | 22/05/2001 | - | - | - | |||
Juveniles | 1 | 8.3 | 04/03/2001 | 10 | 83.3 | 01/06/2001 | 12 | |
1 | 8.3 | 10/04/2001 | - | - | - | |||
Metamórficos | 21 | 2.3 | 07/05/2001 | 878 | 97.6 | 02/05/2001 | 899 | |
Larvas | - | - | - | 550 | 100 | 10/04/2001 | 550 | |
- | - | - | - | - | - | |||
Total | 25 | 1.6 | - | 1465 | 98.3 | - | 1490 | |
2002 | Hembras | - | - | - | 10 | 50.0 | 09/03/2002 | 20 |
- | - | - | 10 | 50.0 | 01/03/2002 | |||
Machos | - | - | - | 10 | 100 | 02/02/2002 | 10 | |
- | - | - | - | - | - | |||
Juveniles | - | - | - | 2 | 100 | 08/06/2002 | 2 | |
- | - | - | - | - | - | |||
Metamórficos | - | - | - | 1523 | 100 | 20/05/2002 | 1523 | |
Larvas | - | - | - | 250 | 44.4 | 14/04/2002 | 562 | |
- | - | - | 312 | 55.5 | 16/04/2002 | |||
Total | 0 | - | - | 2117 | 100 | - | 2117 |
Tabla 3. Avistamientos de individuos de Triturus marmoratus por clase y temporada.
Población | 1ª observación | Nº ejemplares | Ultima observación | Nº ejemplares |
L. Tejar L. Taragabuena L.Vallardon f. Valdematas f. Valdelbuey | 1990 1991 1987 1991 1992 | 25 6 21 6 3 | 1997 1998 2002 2002 2002 | 1 1 0 1 0 |
Tabla 4. Poblaciones reproductoras desaparecidas de Triturus marmoratus en la comarca de Toro
Los censos de peces así como de individuos metamórficos y totales de Triturus marmoratus se observan en las tablas 1, 2 y 3 respectivamente.
Los datos obtenidos reflejan el desajuste que se ha producido en la población norte desde la introducción de Cyprinus carpio en este arroyo, ya que los ejemplares capturados superaban los 500 gramos de peso; lo que implica la depredación masiva de larvas de anfibio. Esto ha reducido el número de larvas y metamórficos drásticamente. En el caso de Rutilus arcasii se cree que siempre ha estado en el lugar, tratándose de una especie autóctona. Según los datos, existía un equilibrio entre Rutilus y la población de Triturus marmoratus (Tabla 2). Dado a que se alimenta principalmente de larvas de dípteros, plecópteros, efemerópteros, ostrácodos, copépodos y ácaros entre otros.
La población sur goza de aislamiento total, ya que no se ha encontrado ningún agente destructor. La fuente no alberga peces, crustáceos ni otra especie de anfibio, lo que facilita una reproducción favorable. Por otro lado no se emplean productos químicos (fitosanitarios) en los huertos colindantes, favoreciendo aún mas la estabilidad reproductora.
El elevado numero de metamórficos censados 4971, no es motivo para decir que la especie es muy abundante. Ya que los jóvenes tritones están expuestos a todo tipo de depredadores (Figura 4), Natrix maura, Ciconia nigra, Vulpes vulpes etc. Se ha estimado una supervivencia anual del 65-79% en adultos (M.García París& JM. Pleguezuelos et al.,2001).
Posibles causas de desaparición de anteriores poblaciones
Hace unos diez años esta especie se extendía por muchas charcas y fuentes de la comarca de Toro (Tabla 4) pero su distribución se ha reducido considerablemente, en parte por el uso indiscriminado de fitosanitarios y el desconocimiento de los agricultores de su correcta utilización, pues recogiendo directamente el agua de charcas y fuentes para la mezcla de producto contaminan los lugares de freza de los anfibios en general. Otra posible causa es la pérdida de ecosistemas idóneos, debido a la extracción de áridos para la construcción. Tierras que en el pasado fueron abandonadas, se han recuperado para la plantación de viñedo, agravando aun más la situación.
Otro factor es la masiva perforación de pozos de sondeo para el riego de tierras de secano, dejando en olvido las balsas tradicionales. Sin olvidar la invasión de especies alóctonas, que están provocando desajustes en todas las poblaciones de anfibios de la zona, siendo Procambarus clarkii el principal destructor de puestas y larvas, y causante de agresiones a adultos (LIZANA Y M. BARBADILLO et al.(1997), A. BERMEJO GARCÍA et al,(2003) (Figura 1).
La Gambusia (Gambusia holbrooki) es otro pececillo introducido para combatir las larvas de mosquito, con un apetito voraz, que afecta sobre todo a puestas y larvas de anfibios (Figura 3). Otro dato relevante es la aparición de la rata común (Rattus norvegicus) en charcas próximas a la ciudad de Toro, atraídas por los cangrejos y peces que se han introducido deliberadamente para volver a capturarlos una vez se reproduzcan. De modo que Triturus marmoratus se ha visto presionado hasta el punto de desaparecer en estos lugares, buen ejemplo es la laguna del Tejar. Donde fue muy numeroso años atrás (Tabla 4).
Las numerosas graveras abandonadas deberían ser rehabilitadas, pues se inundan de agua debido a que la extracción de áridos ha llegado al nivel freático. Esto transforma el lugar en un verdadero humedal, listo para que los anfibios lo colonicen. Hoy en día, los lugares con estas características son empleados como vertederos, contaminando la zona. Podría llegarse a un acuerdo con las empresas mineras, para que cedieran estas zonas al ayuntamiento para que éste rehabilitase el paraje.
La desaparición de especies de zonas que han sido contaminadas o alteradas hace imposible su recuperación, pues estos lugares seguirían estando contaminados en años venideros. Hace años T. marmoratus era una especie abundante, pero los lugares de reproducción de este anfibio se han visto reducidas a dos poblaciones en la zona de estudio siendo, en la actualidad un anfibio en retroceso, que se puede catalogar de amenazado. Sería una lástima perder esta especie en la comarca de Toro, pero la realidad es abrumadora.
Referencias
-LIZANA Y M. BARBADILLO(1997)Legislación y estado de conservación de anfibios y reptiles españoles pp., 477-516.
-ANFIBIOS Y REPTILES DE PENINSULA IBERICA, BALEARES Y CANARIAS(1999)Javier Barbadillo, José Ignacio Lacomba, Valentín Pérez Mellado, Vicente Sancho y Luis Felipe Jurado. pp., 94-96
-ANFIBIOS ESPAÑOLES (2001)Alfredo Salvador y Mario García París. pp., 62-66
- ABEL BERMEJO GARCÍA(2003) Primeros datos de agresiones de Procambarus clarkii sobre Pleurodeles waltl, Butlletí de la Societat Catalana d´Herpetologia 16 pp., 87-91)
AGRADECIMIENTOS
Mª José Bermejo García & Millán Sarmiento por su colaboración en las salidas al campo. Mario García Paris &Miguel Lizana Avía por sus orientaciones.
Campaña de recogida de quelonios alóctonos y autóctonos cautivos en Zamora
Abel Bermejo
C/ Cortes de Toro, 11, 2ºB 49800 Toro (Zamora). España
e-mail: gallipato@herpetologica.org
Keywords: alien species, chelonians, environmental education.
La provincia de Zamora, al igual que el resto de la Península Ibérica , se está viendo afectada por la liberación de diversas especies de quelonios alóctonos. Hasta la fecha, se han hallado dos galápagos exóticos en Zamora, Trachemys scripta elegans y Trachemys scripta scripta. Su reproducción tanto en semilibertad (Martínez Silvestre et al., 1997) como en la naturaleza (de Roa & Roig, 1988, Capalleras & Carretero, 2000, Bertolero & Canicio, 2000) demuestran la adaptación a nuestros humedales. Mingot et al. (2003) ponen de manifiesto la reproducción en la zona centro de la Península Ibérica y Bermejo-García (2006) cita la recolección de siete neonatos de Trachemys s. scripta en las riberas del río Esla. Finalmente, Franch et al. (2006) describen la estructura reproductiva de una población de galápago de Florida en el noreste de la Península Ibérica.
Con estos datos preocupantes, parece lógico impedir su liberación en la naturaleza, con el fin de evitar los problemas que su extracción conlleva. Está demostrado que estos quelonios alóctonos desajustan las cadenas tróficas, afectando directamente a las dos especies autóctonas que pueblan el territorio español, Emys orbicularis y Mauremys leprosa (Barbadillo et al., 1999; Salvador & Pleguezuelos, 2002). Conscientes de esta problemática, surgió la idea de organizar una campaña de recogida de especies exóticas en cautividad. La metodología empleada fue la de informar a la ciudadanía de la provincia mediante carteles informativos y publicaciones en periódicos y revistas locales.
Cabe asimismo mencionar la entrega de quelonios autóctonos retenidos en cautiverio. En efecto, en esta zona es frecuente mantener a Mauremys leprosa y Emys orbicularis en cautividad, ya que según algunos lugareños ahuyentan a roedores. Suelen ser especímenes recolectados en pescas de Procambarus clarkii. Con frecuencia los pescadores retienen a los galápagos junto con los cangrejos durante el transporte, en la creencia que esta acción mantiene con vida a P. clarkii, según comentan los aficionados a esta pesca. Como resultado de esta nefasta práctica, la mayor parte de estos ejemplares sufren agresiones por parte del crustáceo (Figura 1).
Desde 2003 hasta 2006 se han recuperado cuatro galápagos autóctonos (Tabla 1). Estos ejemplares presentaban diversas anomalías debidas al cautiverio: un macho de M. leprosa capturado en una pesca de cangrejos, con extirpación del ojo derecho; una hembra de la misma especie se mantuvo durante más de 5 años en un patio de hormigón presentando terribles callosidades en los miembros posteriores y ausencia parcial de la cola debido a la abrasión con este elemento; un macho joven de E. orbicularis mostraba claramente una fuerte agresión producida por un cánido, este individuo murió pocos días después de la entrega.
Desde el inicio de la campaña se han recolectado 10 ejemplares de galápagos alóctonos (cinco Trachemys scripta elegans y cinco T. s. scripta), hecho que refleja la importancia de este tipo de campañas de recogida que evitan su liberación en nuestros humedales. La mayoría de las personas que donaron uno o varios ejemplares lo hacía por no poder mantener a su galápago. Este problema se hace patente en algunos individuos (n=3) que presentaban anomalías físicas, por falta de espacio y una mala alimentación. Los ejemplares recogidos se han ubicado en instalaciones privadas, zonas creadas especialmente para estos quelonios. También parece razonable, que estas personas tengan derecho de visitar a su antigua mascota si así lo desean. En muchos casos la donación al centro se hizo con esta condición. La adaptación al nuevo entorno incita a estos galápagos a reproducirse bajo estas condiciones (hábitat y alimentación adecuados). Así, se halló una puesta de cuatro huevos en el estanque. Curiosamente fueron localizados en enero de 2007 durante la limpieza del mismo, los huevos se hallaron en el agua. El día 4 de abril de 2008 se observo un neonato de Trachemys scripta elegans, este presentaba diente de huevo y una marcada cicatriz umbilical. La temperatura ambiental en el momento de la observación era de 25º C y las noches anteriores fueron suaves entre 8-10º C, condiciones similares a las expresadas por Albert & Martín (2007) en Alicante, confirmando la reproducción en Zamora bajo condiciones ambientales frías.
Otro aspecto interesante de la campaña es que no se han eliminado estas especies alóctonas como se suele venir haciendo en otras zonas. Se considera que tales ejemplares pueden ser útiles en charlas realizadas durante la visita de centros de enseñanza para concienciar a los jóvenes de la provincia. Generalmente, el motivo de la presencia de estos galápagos exóticos en hogares, se debe a que los niños adquieren la mascota como un juguete. En 2003 se emplearon dos ejemplares de T. s. elegans durante el transcurso de la exposición “Anfibios de la Comarca de Toro” financiada por la Obra Social de Caja Madrid y realizada por Abel Bermejo García, considerándose que esta acción ayudó a los visitantes a reflexionar sobre el respeto y el compromiso que esta fauna y sus problemas merecen.
Agradecimientos: A todas las personas que donaron a su mascota en lugar de liberarlas en la naturaleza. A Manuel Otero Toral por su apoyo incondicional a la causa conservacionista, a Eva Bermejo García y David Olalla Martín que colaboran en la recogida de ejemplares durante la campaña.
Tabla 1. Características de los ejemplares autóctonos recuperados.
Especie | Sexo | Longitud caparazón | Estado | Fecha de entrega |
Mauremys leprosa | Hembra | Callosidades y perdida de cola | 11-011-2003 | |
Mauremys leprosa | Macho | Ojo derecho extirpado | 05-08-2004 | |
Emys orbicularis | Macho | Fuerte agresión de un cánido | 12-10-2004 | |
Emys orbicularis | Macho | Bueno | 16-08-2005 |
Referencias
Albert, A. & Martín, P. 2007. Hibernación post-eclosión de Trachemys scripta elegans en cámara de nido en condiciones naturales en la provincia de Alicante. Bol. Asoc. Herpetol. Esp., 18: 89-90
Barbadillo, L. J., Lacomba, J. I., Pérez-Mellado, V., Sancho, V. & López-Jurado, L. F. 1999. Anfibios y Reptiles de la Península Ibérica ; Baleares y Canarias. Ed. GeoPlaneta. Barcelona.
Bertolero, A. & Canicio, A. 2000. Nueva cita de nidificación en libertad de Trachemys scripta elegans en Cataluña. Bol. Asoc. Herpetol. Esp., 11: 84
Bermejo-García, A. (2006): Impacto de la fauna exótica sobre los anfibios en Castilla y León. Quercus, 239: 30-31
Capalleras, X. & Carretero, M. A. (2000): Evidencia de reproducción con éxito en libertad de Trachemys scripta en la Península Ibérica. Bol. Asoc. Herpetol. Esp, 11(1): 34-35.
De Roa, E. & Roig, J. M. (1998): Puesta en el hábitat natural de la tortuga de Florida(Trachemys scripta elegans) en España. Bol. Asoc. Herpetol. Esp., 9: 48-50
Franch, M.; Llorente, G. A.; & Montori, A. (2006): Ecología reproductiva de Trachemys scripta elegans en el noreste Ibérico. Libro de resúmenes. XIII Congreso Español de Herpetología, San Sebastián: 58.
Martínez- Silvestre, A.; Soler, J.; Solé, R.; González, F. X. & Sampere, X. (1997): Nota sobre la reproducción en condiciones naturales de la tortuga de Florida (Trachemys scripta elegans) en Masquefa (Catalunya, España) Bol. Asoc. Herpetol. Esp., 8: 40-41
Mingot, D.; López-Rodrigo, J.; Ordóñez-Rivas, C. & Sobrino, E. (2003): Reproducción en libertad del galápago de Florida(Trachemys scripta elegans) en el centro de la Península Ibérica. Bol. Asoc. Herpetol. Esp., 14(1-2): 39-43
Salvador, A & Pleguezuelos, J. M (2002) Reptiles Españoles. Canseco Ediciones, S. L., Talavera de la Reina.
MUNIBE (Suplemento/Gehigarria)nº 00 000-000 DONOSTIA-SAN SEBASTIÁN 2003 ISSN XXXX-XXXX MUNIBE (Suplemento/Gehigarria) nº 25-28-33 DONOSTIA-SAN SEBASTIÁN 2007 D.L. SS-1108/07
Efectos de la introducción de especies exóticas
sobre los anfibios de Castilla y León
Effects of exotic species introductions on the amphibians of Castilla y León
ABEL BERMEJO-GARCÍA
C/Cortes de Toro, nº 11, 2B. 48900 Toro (Zamora). España
herpeto31@hotmail.com, gallipato@herpetologica.com
La introducción de especies exóticas es una de las numerosas causas constatadas de alteración de las poblaciones naturales de anfibios. Los peces y crustáceos exóticos introducidos en humedales y depósitos artificiales de agua (fuentes, estanques), que son sistemas cerrados y generalmente de pequeñas dimensiones, constituyen una grave amenaza, tanto para la fase lar-varia como adulta de algunas especies. La competencia ejercida por la fauna invasora empieza a ser documentada en numerosas áreas de la península Ibérica (GALÁN, 1997; LIZANA & BARBADILLO, 1997; RIVERA & SÁEZ, 2003; BERMEJo-GARCÍA, 2003, 2004, 2006A; REBELO et al., 2002; CRUZ & REBELO, 2005; CRUZ et al., 2006).
En la presente nota se abordan los efectos producidos por ciertas especies alóctonas de peces y por cangrejo rojo americano (Procambarus clarkii) en las poblaciones de anfibios de cuatro provincias de Castilla y León (Zamora, Salamanca, Valladolid y León) (figura 1), cuya batracofauna está compuesta por 8 urodelos y 14 anuros(BARBADILLO, 1999; SALVADOR & GARCÍA-PARÍS, 2001;VELASCO et al., 2005). Observaciones antiguas ya detectaron los efectos destructivos de peces invasores sobre las poblaciones locales de anfibios anuros. Tal el caso de la introducción en 1986 de pez gato (Ameiurus melas) en una laguna de Manganeses de la Lampreana (Zamora) (VELASCO et al., 2005), que produjo la desaparición de sus poblaciones de sapillo moteado común (Pelodytes punctatus) y sapillo pintojo (Discoglossus galganoi). En 2002 se iniciaron los muestreos de urodelos en charcas, fuentes y pilones, ante la observación repetida de agresiones de P. clarkii a las colas y extremidades de los adultos de gallipato (Pleurodeles waltl). Los enclaves fueron visitados hasta 2006 con periodicidad mensual. El control de los anfibios afectados se realizó mediante marcaje por amputación de falanges. El muestreo principal se llevó a cabo en la provincia de Zamora, para la que se aportan censos de individuos atacados. En el resto de provincias se recogen observaciones puntuales y se recopilan, en general, los efectos observados por la interacción entre la fauna local y las especies introducidas. Se contó con las observaciones de Pablo García para los enclaves de Salamanca.
RESULTADOS Y DISCUSIÓN
Zamora
El seguimiento durante los últimos cuatro años se ha centrado en poblaciones afectadas por la introducción de P .clarkii, localizadas en el SE y SW de la provincia. El porcentaje de individuos afectados en las tres especies de urodelos muestreados en el conjunto de localidades fue alto en todos los casos, especialmente en Triturus marmoratus. El 79.4 % de ejemplares (n = 68, de cinco localidades) presentaba algún tipo de afección, en todos los casos producida por P. clarkii. El 68.7 % de los individuos de Pleurodeles waltl (n = 265), censados en 10 localidades, presentaba afecciones, esta vez originadas por el cangrejo o ciertos peces igualmente introducidos (Lepomis gibbosus, Gambusia holbrooki y Cyprinus carpio). La especie menos afectada fue Lissotriton boscai, con el 60.8 % de individuos atacados (n = 222, de siete localidades) por P. clarkii y G. holbrooki(tabla I). En alguna localidad, el 100 % de los individuos censados de las dos primeras especies había sido atacado.
Salamanca
La prospección de la región oriental de Salamanca ha servido para poner en antecedentes las causas de regresión en numerosos enclaves de la provincia, generalmente derivadas de la presencia de especies invasoras en los humedales, aunque en un caso pudo comprobarse la extinción de una población de L. boscai por recolecciones de terrariófilos. Además de las especies introducidas en Zamora, en Salamanca se ha detectado la presencia de otros peces, como Gobio lozanoi, Micropterus salmoides y Carassius auratus (tabla II), en charcas, pilones y lagunas de nueva creación. Las introducciones son, en general, producidas por los aficionados a la pesca deportiva y la acuariofilia.
Valladolid y León
En ambas provincias la intensidad de los muestreos ha sido menor, con respecto a las anteriores. En dos manantiales de la zona SW de Valladolid los taxones introducidos fueron peces, que afectaron a la reproducción de T. marmoratus o incidieron en el ciclo larvario de P. waltl, produciendo una severa agresión en las larvas (tabla III). En los dos casos las poblaciones de tritones se encuentran aisladas. En la región SE de León se ha seguido las poblaciones de anfibios en dos lagunas de reciente creación por extracción de áridos, en las que se había introducido P. clarkii. Hasta seis especies de anfibios colonizaron rápidamente las lagunas, pero las poblaciones han sido prácticamente desplazadas por la acción del cangrejo. Los anfibios se ven obligados a reproducirse en otras lagunas, también densamente pobladas por el depredador. Observaciones puntuales realizadas durante el período de estudio ilustran el alcance de los efectos que la introducción de especies exóticas en medios muy limitados producen sobre las poblaciones locales de anfibios. La depredación directa en biotopos artificiales, de reducido tamaño, puede ser evidente, pero sus efectos no menos importantes que si se produjeran en medios naturales, porque los abrevaderos, fuentes y pilones resultan cada vez más importantes para la supervivencia de las poblaciones locales. Al respecto, se comprobó la desaparición de tres especies de anfibios en un pilón de Casas del Conde (Salamanca), por la acción de un solo pez sol introducido en el mismo (P. García, com. pers.). O la de una población de Triturus marmoratus en el arroyo Adalia (Toro, Zamora) por la acción de cinco carpas comunes. Pero la introducción de cangrejos y peces en medios naturales puede resultar un incentivo para que ciertos depredadores de fauna acuática acaben con las poblaciones reproductoras de anfibios, como podría interpretarse la desaparición de todas las puestas de Bufo calamita por la acción de una cigüeña blanca (Ciconia ciconia) en una laguna de Zamora, o la de las diferentes especies de anfibios asentadas en el Tejar (Toro) por la presencia de hasta 20 ardeidas, atraídas por la presencia de cangrejo rojo americano, carpas y pez sol (obs. pers.). Por otra parte, se ha comprobado que el crustáceo, dotado de una gran plasticidad ecológica, seha adaptado sin dificultades al clima local, pudiendo estivar cuando las charcas permanecen secas, sin alimentarse durante periodos de hasta tres meses (obs. pers.). La reactivación del cangrejo con las primeras lluvias otoñales coincide con la freza de algunas especies de anfibios en la zona, que constituirían una importante fuente disponible de alimento en un período crítico para el depredador (BERMEJO-GARCÍA, 2006b).
La contribución real de la fauna exótica al declive de poblaciones de anfibios está todavía por valorar en muchas zonas de Castilla y León, pero los efectos comprobados en las provincias objeto de seguimiento predicen la grave amenaza que para la fauna autóctona suponen las especies invasoras en el conjunto territorial de esta Comunidad Autónoma, donde la práctica de las introducciones es un fenómeno fuertemente instalado en diversos estamentos de la sociedad.
REFERENCIAS
BARBADILLO, L.J., LACOMBA, J.I., PÉREZ-MELLADO, V.,SANCHO, V & LÓPEZ-JURADO, L.F. 1999. Anfibios y reptiles de la península Ibérica, Baleares y Canarias. Barcelona.Editorial Geoplaneta S. A.
BERMEJO-GARCÍA, A. 2003. Primeros datos de agresiones de cangrejo rojo americano (Procambarus clarkii) sobre gallipato (Pleurodeles waltl). Butll. Soc. Cat. Herp., 15: 87-91.
BERMEJO-GARCÍA, A. 2004. Estado de conservación de las poblaciones de Triturus marmoratus en la comarca de Toro (Zamora). Bol. Asoc. Herpetol. Esp., 15(1): 50-53.
BERMEJO-GARCÍA, A. 2006a. Impacto de la fauna exótica sobre los anfibios de Castilla y León. Quercus, 239: 30-31.
BERMEJO-GARCÍA, A. 2006b. Nuevos datos de agresiones de Procambarus clarkii sobre Pleurodeles waltl. Bol. Asoc. Herpetol. Esp., 17(2): 82-85.
CRUZ, M.J. & REBELO, R. 2005. Vulnerability of southwest Iberian amphibians to an introduced crayfish, Procambarus clarkii. Amphibia-Reptilia, 26: 293-303.
CRUZ, M.J., REBELO, R & CRESPO, E.G. 2006. Effects of an introduced crayfish, Procambarus clarkii, on the distribution of south-western Iberian amphibians in their breeding habitats. Ecography, 29: 329-338.
GALÁN, P. 1997. Declive de poblaciones de anfibios en dos embalses de La Coruña (NO de España) por introducción de especies exóticas. Bol. Asoc. Herpetol. Esp., 8: 15-17.
LIZANA, M & BARBADILLO, L.J. 1997. Legislación, protección y estado de conservación de los anfibios y reptiles españo-les, pp. 477-516. In: Distribución y Biogeografía de los Anfibios y Reptiles en España y Portugal. Pleguezuelos, J. M. (Ed.). AHE-Universidad de Granada. Granada.
REBELO, R., CRUZ, M.J., SEGURADO, P., SOUSA, M. & CRES-PO, E.G. 2002. Colapso da comunidade de anfibios da Reserva Natural do Paúl Boquilobo após a introduçao do lagostim-vermelho americano, Procambarus clarkii. VII Congreso Luso-Español de Herpetología, Évora: 78.
RIVERA, X. & SÁEZ, R. 2003. La fauna acuática introducida y su impacto sobre los anfibios y reptiles. Quercus 205: 23-28.
VELASCO, J.C., LIZANA-AVÍA, M., ROMÁN-SANCHO, J., DELIBES DE CASTRO, M. & FERNÁNDEZ-GUTÍERREZ, J. 2005. Guía de los Peces, Anfibios, Reptiles y Mamíferos de Castillay León. Nayalde Editorial. Medina del Campo (Valladolid).
MUNIBE (Suplemento / Gehigarria) 25, 2007
S.C.Aranzadi.Z.E.Donostia/San Sebastián
Bol. Asoc. Herpetol. Esp. (2006) 17 (2)
82
NUEVOS DATOS DE AGRESIONES DE
Procambarus clarkii SOBRE Pleurodeles waltl
ABEL BERMEJO GARCÍA
C/ Cortes de Toro, 11, 2º, B. 49800 Toro. Zamora. España.
Keywords: Alloctonous species, Procambarus clarkii, Pleurodeles waltl.
La provincia de Zamora sé esta viendo afectada por una invasión de cangrejo americano, Procambarus clarkii, en fuentes y pequeñas charcas muy alejadas de ríos y arroyos.
Observaciones propias indican introducciones deliberadas de este cangrejo en estos ecosistemas, con vistas a su captura una vez se reproduzcan. Obviamente, tales introducciones se realizan sin considerar el grave efecto que causa la introducción de esta especie alóctona en poblaciones de anfibios autóctonos. Es bien conocido que el cangrejo americano depreda sobre las puestas y larvas de todas las especies de anfibios. Así pues, Lizana & Barbadillo (1997) citan genéricamente la depredación sobre adultos, en especial sobre tritones; Baila (1992) señala como afecta a Triturus helveticus en el valle del Ebro;Barbadillo et al. (1999) habla de la desaparición de Pleurodeles waltl en balsas donde se ha introducido este invasor y Rebelo et al. (2002) comentan la desaparición de P. waltl, Pelodytes punctatus e Hyla arborea debido a la expansión de P.clarkii. También pueden ocasionar graves daños a adultos de Rana perezi.
MATERIAL Y MÉTODOS
El uno de noviembre de 2002 se inició un censo de una población de P. waltl situada dentro del término municipal de Toro, Zamora (UTM 30TTL99). Se trata de una fuente tradicional, que consta de un aljibe protegido por una bóveda rematada por un arco, el agua sobrante origina una charca. La observación en un elevado número de agresiones sobre ejemplares reproductores, llamó inmediatamente la atención. A partir de esta fecha, en diferentes prospecciones se han venido marcando todos los cangrejos observados en dicha charca, mediante un corte de urópodo (apéndices caudales) a fin de poder evaluar tanto las poblaciones del depredador como de la presa. El marcaje de P. waltl se realiza mediante la amputación de dedos, remarcando una vez se regenera el miembro. En el caso de las larvas, se practicó un corte en la cola. La charca fue dividida en tres sectores A, B, C con una red de malla de plástico, método muy útil para el control de puestas. Los huevos encontrados se contabilizaron por sectores. Se tomaron fotografías de cada ejemplar marcado, asignándoles un código. Se realizaron unas 120 salidas nocturnas, y en todas ellas se observaron agresiones a P. waltl por parte de P. clarkii.
RESULTADOS
La Tabla 1 indica el número de ejemplares de P. waltl censados, marcados y agredidos, tanto en la charca como en el aljibe. En la Tabla 2 se indica el porcentaje de individuos agredidos por temporada y la Tabla 3 muestra los censos de las puestas, larvas e individuos metamórficos durante los dos años que duró el trabajo. El 70% de las puestas censadas fueron depredadas por P. clarki. Del 30% que llegó a eclosionar fueron marcadas 667 larvas, sin embargo, solamente se observaron 6 individuos metamórficos. Se pudo comprobar que los machos suelen presentar mas agresiones, debido a que permanecen en el agua más de 150 días. Los ejemplares marcados como idy12, idy22 idy16 e idy25 estuvieron 175 días en la charca, mientras que las hembras cdx2, cdx7,cdx9 permanecieron menos de 35 días en la charca, siendo por lo tanto menos atacadas.
En la Tabla 4 podemos observar el número de ejemplares de cangrejo americano presente en la zona de estudio. La gran concentración de P. clarkii se debe a su adaptación a este tipo de aguas frías interiores y al éxito reproductor del crustáceo (Bermejo-García, 2006).
DISCUSIÓN
De los resultados se deduce que una considerable proporción de adultos fue atacada y un gran porcentaje de puestas fracasaron debido a la depredación del cangrejo americano. Los ataques se suelen producir en la época reproductora (octubre-marzo), época en la que la concentración de ejemplares de P. waltl, especialmente de machos, en el aljibe y la charca es mayor.
Otro punto interesante respecto a las agresiones (Figura 2), es que se llevan produciendo desde hace tiempo; ya que se ha podido observar partes del cuerpo curadas y en regeneración. Puesto que las agresiones pueden estar repercutiendo en el éxito reproductor de esta especie y teniendo en cuenta que P. waltl es la especie de urodelo más afectada por el cangrejo americano en la Península Ibérica (Cruz &Rebelo, 2005; Cruz et al., 2006), se debe seguir trabajando en este proyecto para valorar realmente el desequilibrio ocasionado por esta especie alóctona. Actualmente se mantienen conversaciones con el Servicio Territorial de Medio Ambiente de la Junta de Castilla y León, para eliminar todos los cangrejos que se encuentren en zonas de freza de esta especie.
2002/03 | En aljibe | En charca | Total | ||||||||
Clase | |||||||||||
EA | % | AA | % AA | ECH | % | ACH | % ACH | ||||
Hembras | 3 | 42,9 | 2 | 66,7 | 4 | 57,1 | 3 | 75,0 | 7 | ||
Machos | 15 | 50,0 | 14 | 93,3 | 15 | 50,0 | 8 | 53,3 | 30 | ||
Total | 18 | 48,8 | 16 | 88,9 | 19 | 51,4 | 11 | 57,7 | 37 | ||
2003/04 | 3 | 27,2 | 2 | 72,73 | 8 | 72,7 | 3 | 37,5 | 11 | ||
Hembras | |||||||||||
Machos | 11 | 57,8 | 10 | 90,9 | 8 | 42,1 | 6 | 75,0 | 19 | ||
Total | 14 | 46,6 | 12 | 85,7 | 16 | 53,3 | 11 | 50,0 | 30 | ||
Tabla 1. Censo de Pleurodeles waltl machos y hembras en aljibe y charca por temporadas. EA: ejemplares marcados en el aljibe; % porcentaje del total de la población censada; AA: ejemplares agredidos en aljibe; % AA: porcentaje de agredidos en aljibe; ECH: ejemplares marcados en la charca; ACH: ejemplares agredidos en la charca; % ACH: porcentajes de agredidos en la charca.
2002/03 Tabla de hembras de Pleurodeles waltl | Tabla de machos de Pleurodeles waltl | ||
Hembras adultas | 7 | Machos adultos | 30 |
Hembras afectadas | 5 | Machos atacados | 22 |
%hembras afectadas | 71,4% | %machos atacados | 73,3% |
2003/04 | 11 | Machos adultos | 19 |
Hembras adultas | |||
Hembras afectadas | 5 | Machos atacados | 16 |
%hembras afectadas | 45,5% | %machos atacados | 84,2% |
Tabla 2: Agresiones en machos y hembras por temporadas.
Censo 2002/03 Puestas/larvas | Sector A | Sector B | Sector C | total | |||
ejemplares | % | ejemplares | % | ejemplares | % | ||
Huevos | 450 | 45,0 | 203 | 20,3 | 347 | 34,7 | 1000 |
Larvas | 120 | 46,7 | 112 | 43,5 | 25 | 9,72 | 257 |
Metamórficos | 0 | 0,0 | 1 | 50,0 | 1 | 50,0 | 2 |
Total | 570 | 38,4 | 316 | 21,2 | 598 | 40,2 | 1259 |
Censo 2003/04 | ejemplares | % | ejemplares | % | ejemplares | % | |
Huevos | 452 | 35,7 | 214 | 16,9 | 600 | 47,3 | 1266 |
Larvas | 201 | 49,0 | 99 | 24,1 | 110 | 26,6 | 410 |
Metamórficos | 2 | 50,0 | 2 | 50,0 | 0 | 0 | 4 |
Total | 665 | 38,9 | 315 | 18,7 | 710 | 42,2 | 1680 |
Tabla 3: Censo de puestas, larvas y metamórficos por temporadas.
2002/03 | aljibe | % | charca | % | total |
edad/sexo | |||||
Hembras adultas | 5 | 62,5 | 3 | 37,5 | 8 |
Machos adultos | 9 | 52,9 | 8 | 47,1 | 17 |
Hembras Jóvenes | 5 | 45,5 | 6 | 54,5 | 11 |
Machos Jóvenes | 4 | 30,8 | 9 | 69,2 | 13 |
Larvas | 18 | 54,5 | 15 | 45,5 | 33 |
Total | 41 | 50,0 | 41 | 50.0 | 82 |
2003/04 | aljibe | % | charca | % | total |
edad/sexo | |||||
Hembras adultas | 14 | 63,6 | 8 | 36,4 | 22 |
Machos adultos | 20 | 51,3 | 19 | 48,7 | 39 |
Hembras Jóvenes | 20 | 57,1 | 15 | 42,9 | 35 |
Machos Jóvenes | 11 | 42,3 | 15 | 57,1 | 26 |
Larvas | 10 | 18,9 | 43 | 81,1 | 53 |
Total | 75 | 42,8 | 100 | 57,1 | 175 |
.Tabla 4: Censo de Procambarus clarkii por temporadas.
Suscribirse a:
Entradas (Atom)